Научно-методические основы оценки и развития специальной аэробной выносливости в академической гребле
Преступая к изложению материала, следует отметить, что
большая часть представленных исследований была проведена в 1977-89 гг. во время
учебы автора в аспирантуре и последующей работы в Ленинградском
научно-исследовательском институте физической культуры (ЛНИИФК), работы в
1978-80 гг. в комплексной научной группе (КНГ) при сборной команде Ленинграда
по академической гребле и 1981-89 гг. в КНГ при сборной команде РСФСР, которую
автор имел честь возглавлять последние 5 лет работы в ней. Также следует
отметить, что при проведении исследований, неоценимую помощь в получении
результатов, их интерпретации и обсуждении оказывали научные сотрудники ЛНИИФКа
и.
Излагая данный, казалось бы, устаревший материал, автор
руководствовался тем, что все новое в спортивной науке, в той части, которая
касается изучения воздействия на организм спортсменов физических нагрузок,
задания интенсивности тренировочных нагрузок, в частности, в академической
гребле - это, по большому счету, хорошо забытое старое. Наши исследования
дополнены обзором и критическим анализом современной литературы по
медико-биологическим вопросам подготовки в видах спорта с преимущественным
проявлением выносливости. Наблюдения за тренировочным процессом членов сборной
команды России по академической гребле на данном временном этапе, за уровнем их
специальной подготовленности, который не соответствует мировому, беседы с
тренерами - убедили нас в актуальности представленного материала.
Если говорить о циклических видах спорта, то к традиционным факторам, определяющим спортивную работоспособность, относят максимальное потребление кислорода (МПК, Vo₂max), порог анаэробного обмена (ПАНО) и экономичность выполнения соревновательного движения (бега, плавания, гребли), которая в большой степени зависит от ритмичного чередования напряжения и расслабления мышц, совершающих его. МПК отражает потенциал организма вырабатывать энергию путём аэробного метаболизма. Аэробный метаболизм является более эффективным путем выработки энергии, нежели анаэробный (бескислородный), хотя оба между собой тесно взаимосвязаны. Максимальная величина этого показателя зависит от способности лёгких и кровеносной системы транспортировать кислород, а мышц его использовать.
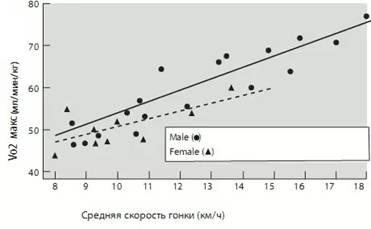
Рис. 1. Пропорциональная зависимость работоспособности (скорости бега на марафонской дистанции) от Vo₂max (МПК) (С. Илюков, 2011)
В последнее время большее распространение находит
представление максимального потребления кислорода по аллометрическому методу,
где учитывается строение и состав тела. Этот метод гораздо более точен при
долгосрочном наблюдении за развитием аэробных способностей спортсмена, когда со
временем меняется как состав тела, так и его конституция.
Величина максимального потребления кислорода зависит от развитости системы связывания, переноса и использования кислорода, которая, в свою очередь, состоит из ряда звеньев.
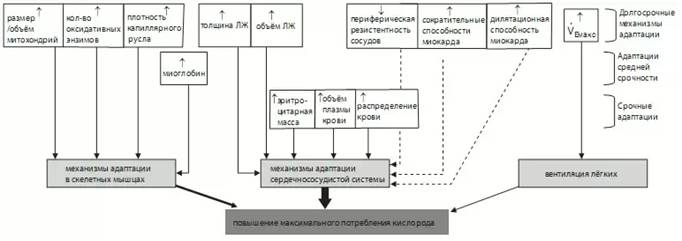
Рис. 2. Схематичное изображение звеньев переноса и потребления кислорода в организме (С. Илюков, 2011)
Основной функцией сердечно-сосудистой и дыхательной систем является поддержание клеточного дыхания. Слаженность взаимодействия функционирования этих систем отражена в потреблении кислорода (О₂) и выделении углекислоты (СО₂) в ответ на определенный уровень работы и их отношение к частоте сердечных сокращений, вентиляции и друг к другу.
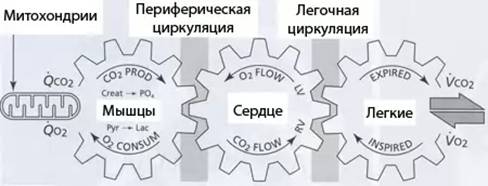
Рис. 3. Схема газообмена у человека (2007)
Условно цепочку транспорта кислорода можно поделить на
центральный и периферический компоненты. К центральному относятся лёгкие,
сердце и кровеносная система, а к периферическому отделу следует относить ткань
поперечнополосатой мускулатуры.
В центральной части в свою очередь отдельно выделяют: поперечное сечение и объём стенки левого желудочка (ЛЖ), дилятационные способности миокарда, объёмы плазмы крови и массы кровяных телец. Левый желудочек образован более толстыми мышечными волокнами, чем правый, так как он противостоит более высокому давлению крови в большом круге кровообращения и должен совершать большую работу по его преодолению во время систолы. Толщина стенки правого желудочка колеблется от 2 до 6 мм, а левого – от 10 до 14 мм.
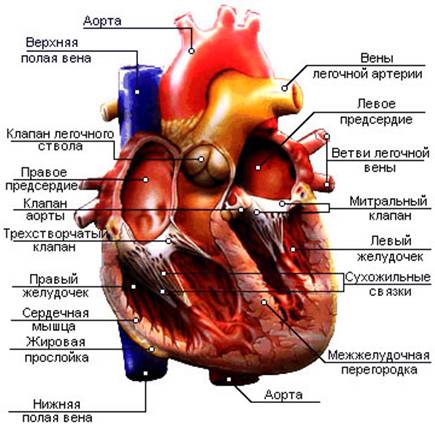
Рис. 4. Строение сердца
В периферической части выделяют: плотность капиллярного
русла, количество и соотношение мышечных волокон разного типа, объём
митохондриальных, оксидативных энзимов и концентрацию миоглобина.
Митохондрии (отдельные и множественные) это субклеточные структуры, содержащиеся во всех клетках, в которых осуществляются реакции цикла Кребса и происходит перенос электронов. Цикл Кребса - это последовательность химических реакций, происходящих в митохондриях, в результате которых вырабатывается двуокись углерода и происходит ионизация атомов углерода – ионы водорода и электроны отщепляются от атомов. Этот процесс называется циклом трикарбоновых кислот (ТСА) или циклом лимонной кислоты.
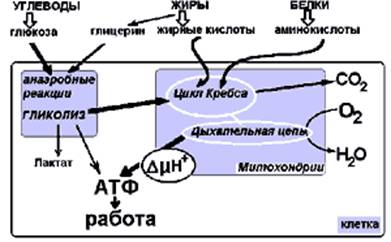
Рис. 5. Схема окисления углеводов, жиров и белков в реакциях энергетического обмена.
Реакции гликолиза идут анаэробно (без кислорода) и
завершаются накоплением лактата. В аэробных условиях продукты гликолиза,
окисления жирных кислот и аминокислоты сгорают - окисляются с участием
кислорода в митохондриях - клеточных органеллах, выполняющих роль основного
источника энергии. Аккумуляция энергии в виде трансмембранного потенциала и АТФ
(аденозинтрифосфорной кислоты) происходит в дыхательной цепи - полиферментном
комплексе, расположенном в мембране митохондрий.
Митохондрии, которые в этом процессе забирают кислород, представляют собой энергетическую фабрику клетки и часто называются "аэробная печка". Это название они получили из-за того, что кислород и топливо соединяются в процессе выделения энергии, результатом которого является образование АТФ, которая присутствует во всех мышечных клетках. Клетка может выполнять работу только в результате выделения энергии, происходящего при разложении АТФ.
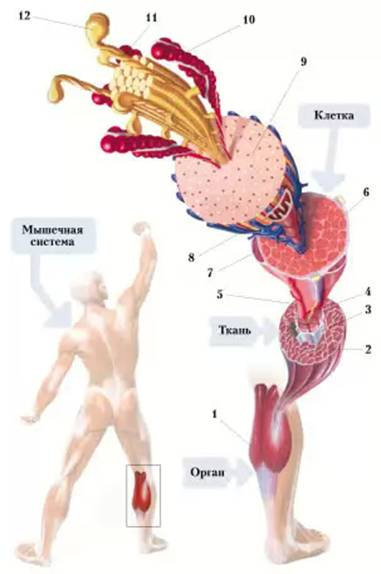
Рис. 6. Система – орган – ткань – клетка:
1 - орган (икроножная мышца); 2 - поперечно-полосатая мышечная ткань (срез); 3 - пучок мышечных волокон; 4 - двигательный нерв; 5 – капиллярная сеть; 6 – многоядерная мышечная клетка (волокно); 7 – одно из ядер мышечной клетки; 8 – митохондрия; 9 – миофибрилла; 10 – актин; 11 – миозин; 12 – миозиновая головка.
Чем больше митохондрий у спортсмена, тем выше его
выносливость. Причина в том, что это единственные клетки, в которых углеводы,
жиры и протеины могут распадаться в присутствии кислорода, выделяя энергию для
двигательного действия. Таким образом, суть тренировочного процесса в видах
спорта с преимущественным проявлением выносливости заключается в увеличении
плотности митохондрий и увеличении концентрации цитохрома С, вещества
находящегося внутри митохондрий, которое имеет огромное значение для выделения
энергии в результате аэробных процессов.
Митохондрии обладают собственной генетикой, и все
митохондрии в теле человека унаследованы от матери. Это происходит из-за того,
что женская яйцеклетка имеет митохондрии, а у сперматозоидов их нет. Несмотря
на широко распространенное мнение, способность к высоким спортивным достижениям
наследуется от матерей, а не от отцов.
Процесс увеличения митохондрий можно описать следующим
образом: во время тренировки происходит энергетическое истощение мышечных
клеток, которое вызывает образование медиаторов, воздействующих на генетический
аппарат мышечных клеток. Генетический аппарат при этом активизируется, запуская
белковосинтетические процессы (протеинсинтез). Активация протеинсинтеза ведет к
гипертрофии митохондрий и увеличению их количества. Если рассмотреть под
микроскопом мышечные клетки стайеров, мы сможем увидеть много крупных и
«развитых» митохондрий, большое количество которых находят в таких частях тела
как сердце, эндокринных железах и печени.
Далее приводятся данные из статьи Фрэнка Хорвилла (Frank
Horwill)-легендарного английского тренера (умер 1 января 2012 г. в возрасте 84
лет), широко известного в мировых легкоатлетических кругах, тренировавшего
спортсменов Великобритании, Канады, Зимбабве, Ирландии, Польши, Кении,
Бахрейна, Португалии и Южной Африки «Что вы должны знать о митохондриях и как
это может изменить вашу тренировку».
Первые попытки физиологов увеличить плотность митохондрий,
осуществлялись через воздействие на эндокринную систему, и эти попытки имели
определённый успех. Количество митохондрий возрастало вместе с ростом уровня
гормона щитовидной железы - тироксина. Лабораторным крысам добавляли в пищу
высушенную щитовидную железу, и это приводило к существенному росту размеров и
плотности митохондрий в печени и в сердце. В качестве стимулирующего препарата
тироксин был очень популярен в течение определённого времени, пока не было
обнаружено, что избыточная концентрация этого гормона приводит к весьма
нежелательным побочным эффектам.
Джон Холосци, физиолог из Медицинской Школы Вашингтонского
Университета в Сен Луисе обнаружил, что продолжительные упражнения приводят к
увеличению количества митохондрий. Он заставлял одну группу лабораторных крыс
бегать на тредбане по 2 часа в день с интенсивностью% от МПК в течение 12
недель, в то время как другая группа сидела безвылазно в клетках. В конце
тестового периода Холосци обнаружил, что крысы за время выполнения упражнений
увеличили количество митохондрий на%, и кроме того у них удвоилась концентрация
цитохрома С. Работы Холосци подтвердили, что методика увеличения выносливости
Ван Аакена посредством продолжительного медленного бега была совершенно
обоснованной. Холосци продолжал свои исследования. Теперь одна группа мышей
бежала в течение 10 минут в день, другая 30 минут, третья 60 минут в день и
четвёртая в течение 2-х часов. Тренировка проводилась пять дней в неделю в
течение 13 недель со скоростью примерно% от МПК для тренированной лабораторно
крысы. Как и ожидалось, те, кто бегали 2 часа в день имели самое хорошее
развитие митохондрий. Занимавшиеся 10 минут в день увеличили количество
цитхрома С на 16% больше по сравнению с группой нетренированных крыс, бегавшие
30 минут имели увеличение на 31%, бегавшие час - на 38% и бегавшие 2 часа - на
92%. В 1967 году эти результаты стали сильнейшим аргументом в поддержку
методики длительного медленного бега. Связь между работоспособностью и
количеством цитохрома С была твёрдо установлена.
Исследования Холосци с ликованием были приняты
последователями методики А. Лидьярда. Его программа заключалась в постепенном
увеличении объёма медленного бега до 100 миль в неделю, в течение 10 недель
зимнего периода. Некоторые бегуны, например, Дэвид Бедфорд доводили
тренировочный объём до 200 миль в неделю, выполняя 3 тренировки в день. Однако
у работы Холосци, при всех её достоинствах был существенный недостаток - она
ничего не говорила о влиянии интенсивности бега на развитие митохондрий - все
крысы бегали с одинаковой скоростью.
Гари Дадли, сотрудник Нью Йоркского государственного
университета в Сиракузах, в 1982 году исследовал влияние интенсивности
выполняемой работы на рост числа митохондрий. Он провёл очень кропотливую
работу - крысы бегали пять дней в неделю от 5 до 90 минут в день, в течение 8
недель. Интенсивность бега варьировалась от 40% до 100% МПК. Дадли также провел
пионерское исследование, определяющее зависимость воздействия скорости и
продолжительности упражнений на развитие различных мышечных волокон (быстрые
волокна, аэробные быстрые волокна, медленные волокна). Результаты оказались
следующими.
Тренировка продолжительностью более 60 минут не оказывала
влияния на увеличение количества цитохрома С. Тренировка продолжительностью от
30 до 60 минут, приводила к увеличению цитохрома С, но продолжение работы более
60 минут (в промежутке от 60 до 90 минут) его уровень уже не поднимала. Это
наблюдение было верным для работы любой интенсивности, которые исследовал
Дадли, и для всех типов мышечных волокон. Развитие митохондрий прекращалось
через час.
Тренировавшиеся по 10 минут в день с интенсивностью 100% от
МПК подняли концентрацию цитохрома С в 3 раза.
Бег в течение 27 минут с интенсивностью 85% от МПК поднял
содержание цитохрома С на 80%.
Тренировка продолжительностью от 60 до 90 минут с
интенсивностью от 70% до 75% от МПК (марафонская скорость), подняла содержание
цитохрома С только на 74 процента.
В промежуточных мышечных волокнах (по своим характеристикам,
стоящим между медленными и быстрыми) было зафиксировано сходное влияние
интенсивности. Десять минут ежедневного быстрого бега приводило к такому же
увеличению количества цитохрома С, что и 27 минут бега с интенсивностью 85% т
МПК или от 60 до 90 минут бега с интенсивностью от 70% до 75% от МПК.
Лучшей стратегией для увеличения количества цитохрома С в
медленных волокнах оказался 60 минутный бег с интенсивностью 70% - 75% от МПК.
Это приблизительно от 80% до 84% от максимального пульса. Такая тренировка
поднимает количество цитохрома С на 40%.
Бег с интенсивностью 85% от МПК, в течение 27 минут привёл к
повышению цитохрома С на 28%.
Быстрый бег с интенсивностью 100% от МПК, поднимает
содержание цитохрома С в медленных волокнах на 10%, что не удивительно, так как
медленные волокна задействованы во время интенсивного бега в гораздо меньшей
степени чем быстрые. Однако, бег с данной скоростью поднимает содержание
цитохрома С на 1% в минуту.
Далее, 90 - минутный бег с интенсивностью от 70 до 75% от
МПК поднимает содержание цитохрома С на 0,75% в минуту (Frank Horwill).
При выполнении физической работы задействованы три системы: кровь, сердце и мышцы. В крови лимитирующим моментом может быть кислородная емкость крови. Этот фактор зависит от общего объема крови и содержания в ней красных кровяных телец, содержащих гемоглобин, который и является носителем кислорода в крови.

Рис. 7. Величины максимального потребления кислорода в зависимости от величины ударного объема сердца (Т. Нильсен,1990).
Третьим лимитирующим фактором могут быть мышцы. Для их
развития необходимо использовать различные виды тренировочных программ, как на
длинных дистанциях, так и интервальные тренировки. Работая на длинных
дистанциях, у спортсмена развиваются мышечные ткани, капиллярная плотность
мышц, увеличивается энзимная активность, увеличивается число
митохондрий в клетке. Все это позволяет увеличить окислительную способность
организма, повысить уровень порогов аэробного и анаэробного обменов.
Интервальные тренировки способствуют увеличению ударного объема сердца,
улучшают сердечно – сосудистую систему.
Величина максимального потребления кислорода (Vo₂max) –
интегральный показатель, характеризующий суммарную мощность как аэробных, так
анаэробных систем энергообеспечения во время максимальной физической нагрузки.
Vo₂max у нетренированных мужчин (25-30 лет) составляет в среднем 40-45
мл/кг/мин. При регулярных тренировках Vo₂max увеличивается до 50-55 мл/кг/мин.
У элитных спортсменов в видах спорта с преимущественным
проявлением выносливости Vo₂max превышает 80 мл/кг/мин. У Бьерна Дэли (Bjorn
Daehlie) имеется самый высокий уровень Vo₂max, равный 90 мл/кг/мин. из всех
лыжников-гонщиков. Только еще два спортсмена в мире имеют величину Vo₂max,
превышающую 90 мл/кг/мин., к ним относятся Индурайн (велоспорт) и Морселли (бег
на средние дистанции).
Хотя следует отметить, что наивысшая зарегистрированная
величина МПК - 94,0 мл/кг/мин – была у экс - рекордсмена мира в беге на 5000м Эмиля
Путтеманса, Бельгия. У рекордсмена России в беге на 5000 м, этот рекорд до сих
пор еще не улучшен) Валерия Абрамова величина МПК - 77, 0 мл/кг/мин, т. е.
значительно меньше, а спортивный результат был лучше. Для сравнения этих
значений можно привести данные о том, что скаковая лошадь имеет Vo₂max, равное
свыше 150 мл/кг/мин.
Основываясь на многочисленных данных, можно утверждать, что
потолок Vo₂max достигается за 6-8 лет тренировок и в большой степени результат
зависит от наследственной предрасположенности к видам спорта с преимущественным
проявлением выносливости, а также подчёркивает актуальность исследований в
области спортивной генетики и использования этих результатов в спортивной
селекции на ранних стадиях подготовки спортсменов. К генетическим факторам,
лимитирующим достижения в видах спорта с преимущественным проявлением
выносливости, можно отнести также ширину устья аорты. Чем шире аорта у
спортсмена, тем с меньшим напряжением происходит работа его сердца при
интенсивной двигательной деятельности. Оптимальные величины ширины устья аорты
у гребцов, по данным (1983), у женщин 3,1-3,7 см, у мужчин - 3,5-4,2 см. В
качестве примера представим данные МСМК по академической гребле Александра
Устинова. При росте 186 см и весе 82 кг ширина устья аорты, измеренная методом
ЭхоКГ составляла у него 4,5 см. У ЗМС Галины Ермолаевой, неоднократной
чемпионки мира, серебряного призера Олимпийских игр 1976 г. по академической
гребле -4,5 см, а у ЗМС В. Парфеновича, неоднократного чемпиона мира и
Олимпийских игр 1980 г. по гребле на байдарке – 5 см.
Представляет интерес данные длительного эксперимента, в
котором на протяжении 6,5 лет наблюдали за изменением показателей физической
работоспособности у финских лыжников уровня молодежной сборной в сравнении со
сверстниками из Норвегии (С. Илюков, 2011). Наблюдение
началось, когда исследуемым было в среднем по 16 лет, а по завершении, их средний возраст составил 22 года. В ходе эксперимента выяснилось, что прирост работоспособности идёт как за счёт развития центральных, так и периферических звеньев системы транспорта кислорода. При этом полости сердечной мышцы (важный компонент, который определяет, сколько крови сердечная мышца будет способна прокачивать за одно сокращение) развивались и увеличивались в первые три года наблюдения, в возрастном промежутке от 16 до 19 лет, после чего сердечная мышца стала развиваться за счет увеличения поперечного сечения, которое влияет на силу сокращений миокарда.
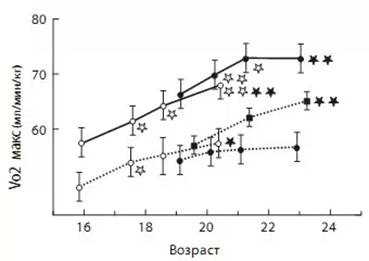
Рис.8. Динамика Vo₂max (МПК) в ходе проведения эксперимента
Под конец эксперимента у части лыжников рост МПК(Vo₂max)
выровнялся и вышел на плато и вместе с тем затормозился прирост показателей сердечно-сосудистой
системы. Один из интересных фактов, отмеченных в исследовании, заключался в
том, что те лыжники, у которых показатели работоспособности (объёмы полостей
сердца, Vo₂max и т. д.) были достаточно высокими в 16 летнем возрасте,
продолжили пропорциональный рост и в дальнейшем, по-прежнему обгоняя своих
сверстников. Те спортсмены, которые отставали по средним показателям в раннем
возрасте, сохранили эту разницу и на более позднем этапе. Это ещё раз
подчеркивает необходимость целенаправленного поиска талантов и отбора в спорте.
Высокий уровень максимального потребления кислорода является
одним из предпосылок достижения спортсменом высокого соревновательного уровня,
однако не предопределяет его безоговорочную успешность. Эта закономерность
особенно хорошо видна среди элитных спортсменов с высокими показателями
максимального потребления кислорода, но существенной разницей в спортивных
результатах.
Выход Vo₂max на плато с годами систематических тренировок отмечен во многих видах спорта с преимущественным проявлением выносливости. В исследовании Мартина с высококвалифицированными американскими бегунами в подготовке к Олимпийским играм в течение 2,5 лет не было никаких сдвигов в показателях МПК. Несмотря на это, регистрировался постоянный регулярный рост спортивных результатов. На частном примере обладателя мирового рекорда в марафонском беге среди женщин Полы Рэдклифф (Paula Radcliff) видно (рис. 5), что она достигла потолка максимального потребления кислорода в 70 мл/мин/кг будучи в 18 летнем возрасте, после чего её спортивная работоспособность повышалась за счёт развития других качеств.
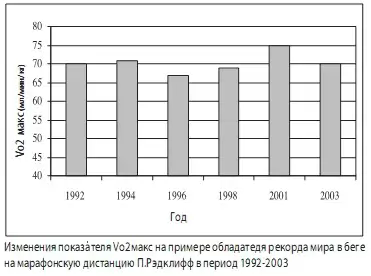
Рис. 9. Изменение Vo₂max обладателя рекорда мира в беге на марафонскую дистанцию среди женщин Полы Рэдклифф в период гг. (A.M. Jones, 2006)
A. Legaz Arrese et al. (2005) проводили исследования на
большой группе (33) квалифицированных бегунов в течение трех лет. Результаты
возросли в среднем на 1.77% у мужчин и на 0.69% у женщин, в то время как
значения Vo₂max (МПК) практически не изменились (76.56 – 76.42) у мужчин и
(70.31 – 70.05) у женщин, что показывает независимость спортивного показателя и
уровня Vo₂max. Таким образом, в однородных группах, таких как хорошо
подготовленные стайеры, Vo₂max не коррелирует со спортивным результатом и не
может характеризовать сильнейших спортсменов (A. Legaz Arrese et al., 2007).
В исследованиях F. Scharhag-Rosenberger et al. (2009)
установлено, что при тренировке с интенсивностью на одинаковом проценте от
Vo₂max были достигнуты различные метаболические сдвиги. Было определено, что
существует значительный индивидуальный разброс в показателях лактата в ответ на
одинаковую интенсивность даже, если участники эксперимента имели одинаковый
уровень Vo₂max. Это привело авторов к заключению, что если мы желаем произвести
аналогичные сдвиги метаболизма, то значение Vo₂max не должно учитываться. В
дополнение к различным уровням лактата, другие факторы, такие как тип мышечных
волокон и иные физиологические параметры варьируют при одинаковых значениях
Vo₂max. N. B.J. Vollard et al. (2009) выявили, что если в среднем для всей
группы были достигнуты сдвиги в параметрах выносливости после шести недель
тренировки с интенсивностью на уровне 70% от Vo₂max, то рассматривая сдвиги
индивидуальных параметров можно было отметить широкий спектр разнообразия.
Таким образом, базовая тренировка в видах спорта с
преимущественным проявлением выносливости, основанная на скорости преодоления
дистанции на уровне или в процентах от скорости на уровне Vo₂max (МПК),
приводит к широкому спектру индивидуальных реакций даже среди однородных групп
и не является «волшебным стимулом» воздействия на организм спортсменов, каким
иногда его представляют (S. Magness, 2009). Например, порог анаэробного обмена
(ПАНО) может находиться при разных процентах от Vo₂max даже у подготовленных
спортсменов (G. A. Brooks et al., 2004) Эти исследования еще раз подтверждают
вывод о том, что для подготовленных спортсменов нет смысла проводить
специальную тренировку на совершенствование уровня Vo₂max. Как пример, можно
показать, что если два спортсмена выполняют нагрузку при 80% от Vo₂max, то один
делает это до уровня порога анаэробного обмена, а другой выше. Это может
существенно влиять на энергетику тренировочной работы. Существенное увеличение
показателя Vo₂max (МПК) отмечается только на начальном этапе подготовки
спортсменов.
Управление тренировочным процессом спортсменов невозможно
без установления количественных критериев и зависимостей, связывающих такие
важные параметры физической нагрузки как объем и интенсивность применяемых
средств с ответной реакцией организма. Результаты выступлений гребцов в
соревнованиях зависят от гармоничного развития всех физических качеств при
ведущем значении специальной силовой и общей выносливости. Создание
функциональной базы выносливости в подготовительном периоде осуществляется
средствами общей физической подготовки (ОФП) и специальной физической
подготовки (СФП) за счет упражнений, выполняемых в режиме, постепенно
приближающемся к предельным возможностям организма работать в аэробных условиях
энергообеспечения (, 1973 и др.)
Результатами исследований в циклических видах спорта
установлена эффективность совершенствования выносливости при использовании
продолжительных нагрузок, которые по воздействию на организм не уступали бы
соревновательной деятельности (1971, 1975, 1971 и др.). В подготовительном
периоде наибольшие сдвиги в росте данного качества оказывают тренировочные
нагрузки на длинных отрезках, преодолеваемых со скоростью 80-90% от планируемой
средней соревновательной (1973, 1974, 1975 и др.). Менее длинные отрезки
дистанции целесообразно преодолевать со скоростью не более, чем на 8-10%
превышающую среднюю соревновательную (1971 и др.).
В циклических видах спорта объем тренировочной работы
учитывается по времени физической нагрузки или пройденному пути, задание же
интенсивности представляет определенную сложность. В практике академической
гребли ее, как правило, задают темпом гребли и ЧСС, что, по нашему мнению, в
большинстве случаев не оправдано. Мы полагаем, что лидирование по скорости
передвижения, тесно связанной с мощностью выполнения двигательной деятельности,
при одновременном наблюдении за изменением темпа и ритма движений, ЧСС,
является более рациональным методом дозирования и оперативного контроля
временных характеристик воздействия нагрузки заданной интенсивности. При общей
физической подготовке лидирование производится по времени преодоления стандартных
отрезков, а в гребле - с сопровождающего лодку катера, на котором установлен
датчик скорости, GPS-навигатор и цифровой анемометр, позволяющий следить за
изменениями скорости и направления воздушного потока. При этом ориентиром
скорости специальной тренировочной нагрузки должна быть, прежде всего, средняя
соревновательная скорость планируемого результата (в процентах от нее) и
уровень максимальной скорости на данный этап времени.
Целесообразность данного подхода к дозированию временных
характеристик нагрузки в академической гребле была нами проверена в ходе
исследования (1984, 1985), задачей которого являлось изучение воздействия на
организм спортсменов физической нагрузки постоянной мощности при индивидуально
дозированной ее длительности. Критерием окончания воздействия нагрузки служило
начало снижения ЧСС, что свидетельствует об адаптации сердечно-сосудистой за
счет увеличения пульсового артериального давления (H. Shibayama, H. Ebashi,
1980). Для решения поставленной задачи произведено обследование 8-х гребцов в
возрасте 16 лет. Нагрузкой служила работа на велоэргометре ЕМ-369 фирмы
Elema-Schönander, индивидуальной для каждого испытуемого мощности при ЧСС 150
уд/мин. До нагрузки, через 20 минут, 6 и 24 часа после ее окончания определялся
показатель PWC₁₇₀.
Во время работы регистрировалась ЧСС, измерялось
артериальное давление и определялись в капиллярной крови следующие биохимические
показатели: лактат по Штрому, гемоглобинцианидным методом гемоглобин,
гематокрит, кислотно-щелочное равновесие методом Аструпа и методом
конкурентного связывания кортизол. Длительность индивидуально дозированной
физической нагрузки составляла по группе от 82 до 87 минут, задаваемая мощность
от 1100 до 1450 кгм/мин, максимальная ЧСС к моменту окончания воздействия от
177 до 192 уд/мин.
Изменение регистрируемых и определяемых показателей по одному из спортсменов представлено в таблице 1.
Таблица 1
Динамика воздействия физической нагрузки постоянной мощности
|
Показатели |
Исходные данные |
Время нагрузки, мин |
|||||
|
12 |
28 |
60 |
68 |
82 |
87 |
||
|
ЧСС, уд/мин |
80 |
162 |
171 |
181 |
182 |
192 |
188 |
|
АД, мм. рт. ст. |
130/70 |
165/50 |
- |
- |
- |
170/30 |
170/20 |
|
Лактат, мМ |
2.3 |
3.4 |
2.5 |
2.4 |
2.4 |
2.4 |
2.2 |
|
Hb, г,% |
15,3 |
13,9 |
15,0 |
15,0 |
15,3 |
13,3 |
13,3 |
|
Гематокрит, % |
44 |
- |
- |
- |
- |
- |
40 |
|
pH |
7,48 |
- |
7,42 |
7,46 |
7,47 |
7,45 |
7,43 |
|
BE, мэкв/л |
+ 4 |
- |
0 |
-1 |
-1,5 |
-2 |
-1,5 |
|
pCO₂, мм. рт. ст. |
40 |
- |
38 |
36,5 |
35,5 |
38,2 |
35,0 |
|
ВВ, мэкв/л |
45 |
- |
40 |
41 |
42 |
42 |
44 |
|
SB, мэкв/л |
29 |
- |
25 |
26 |
26 |
26 |
23,5 |
|
Кортизол, нг/л |
79,6 |
- |
74,4 |
84,8 |
- |
116,4 |
120,5 |
Анализ биохимических сдвигов позволил установить, что
несмотря на значительное увеличение ЧСС, нагрузка выполнялась в аэробной зоне
энергообеспечения. Снижение ЧСС к концу нагрузки свидетельствует, по-видимому,
об адаптации организма спортсменов к данному воздействию за счет увеличения
пульсового артериального давления. Показатель PWC₁₇₀ через 20 минут после
окончания нагрузки снижался по группе до 72,9-91,5 % от исходного значения и
возрастал до 112-116% через 24 часа.
Характерной особенностью построения системы подготовки на
этапе спортивного совершенствования является преимущественное использование
узкого круга специализированных средств тренировки, а также управление
тренировочным процессом с ориентацией на модельные характеристики соревновательной
деятельности (1982, 1995).
Основу тренировочных программ, направленных на создание
фундамента специальной подготовленности, в видах спорта с преимущественным
проявлением выносливости, должны составлять нагрузки на уровне порога
анаэробного обмена (ПАНО или как его еще обозначают АнП, или англ. AT -
anaerobic threshold), доля которых должна быть более 50% от общего годичного
объема и силовая подготовка, являющаяся адекватным стимулом развития рабочей
гипертрофии мышц.
Этот фундамент у высококвалифицированных спортсменов
характеризуется высоким значением МПК, высокой скоростью передвижения на уровне
ПАНО (в пределах 85-95% от скорости передвижения при МПК), эффективной и
экономичной спортивной техникой, сглаживанием реципрокных отношений между
жировым и углеводным обменом, высоким специфическим уровнем силы мышц,
обеспечивающих двигательное соревновательное действие (1987 и др.).
Многие годы вполне обоснованно усиленной критике
подвергается применяемое в практике положение о задании интенсивности
физических нагрузок при использовании усредненных критериев образования
молочной кислоты(лактата) в крови на уровне 2 или 4 мМоль/л как показателей,
характеризующих соответственно порог аэробного и анаэробного обмена (1982; K. Tanaka
et al., 1983; S. Aunola, *****sko, 1984, 1985 и др.). Это обусловлено тем, что
большая функциональных показателей у человека имеет высокую межиндивидуальную изменчивость,
коэффициент вариации, которой достигает 30% по сравнению с 3-7% анатомических
признаков (1980 и др.). Причем, относительная вариабельность увеличивается от
антропометрических показателей к физиологическим и далее к биохимическим (1987
и др.). Следовательно, для оценки состояния человека необходимо устанавливать
индивидуальные функциональные нормы и не пользоваться усредненными данными (1984)
В связи с вышеизложенным не выдерживает критики концепция деления интенсивности
тренировочных нагрузок по следующим зонам: концентрация лактата в крови до 4
мМоль/л-работа аэробной направленности; до 8 мМоль/л-работа при смешанном
(аэробно-анаэробном) энергообеспечении; свыше 8 мМоль/л-анаэробное
гликолитическое энергообеспечение, - выдвигаемое большим количеством авторов.
Особо следует отметить вариабельность уровня молочной
кислоты в крови в состоянии покоя. По мнению и (1983), повышенная концентрации
ее утром (более 2мМоль/л) и сниженный уровень pH (по сравнению с 7,40 ± 0,05)
свидетельствует о выраженном недовосстановлении. Мы же наблюдали и большие
значения лактата (без определения рH крови) при субъективно хорошей самооценке
состояния квалифицированными спортсменами.
Подводя некоторый итог, можно утверждать, что интенсивность
большинства развивающих функции организма тренировочных нагрузок, используемых
в циклических видах спорта с преимущественным проявлением выносливости, должна
находится в диапазоне от скорости (или мощности) на уровне порога анаэробного
обмена (ПАНО) до скорости на уровне максимального потребления кислорода
(Vo₂max). Это связано с тем, что при физической нагрузке ниже ПАНО (т. е. до
развития лактат-ацидоза) реакции дыхания и кровообращения сохраняют pH, pCO₂ и
pO₂ артериальной крови на уровне, близком к стационарному состоянию в условиях
покоя (B. J. Whipp, S. A. Ward, 1982 и др.). Тренировочные воздействия,
мощность двигательного эффекта которых превышает критическую (т. е. мощность на
уровне Vo₂max), способствуют развитию анаэробных механизмов энергообеспечения и
используются в данных видах спорта в гораздо меньшем объеме и главным образом в
соревновательном периоде. Эти положения установлены и подтверждены в
методически правильно выполненных, по нашему мнению, отечественных исследованиях
(1973, 1982, 1983, 1985, 1986, 1987; К. Сахновский, С. Фесенко, 1987 и др.) и в
большом количестве работ, опубликованных за рубежом.
Следует особо отметить отличие локализации порога
анаэробного обмена при различных тестирующих эргометрических нагрузках (J. A.
Davis et al., 1976; R. T. Withers et al, 1981 и др.) Это подтверждает тот факт.
что ПАНО отражает уровень функциональных возможностей при выполнении конкретной
физической работы и зависит от мышечной массы, участвующей в данных
двигательных действиях (1983; K. Wasserman, 1984).
Несмотря на то, что ПАНО можно классифицировать как критерий
аэробных возможностей, его изменения нельзя полностью связывать с динамикой
максимального потребления кислорода, т. к. установлено, что уровень порога
анаэробного обмена может меняться при неизменном Vo₂max, а выполнение
упражнений при потреблении кислорода 50-85% от Vo₂max приводит к разному
рабочему стрессу у испытуемых с различным уровнем ПАНО, но с одинаковой величиной
МПК (J. Dwyer, bee, 1981).
Таким образом, можно констатировать, что порог анаэробного
обмена, представляющий переходный момент в метаболическом стрессе, является
лучшей основой для планирования интенсивности тренировочных воздействий в видах
спорта с преимущественным проявлением выносливости, чем максимальное
потребление кислорода (V. Bung, 1983; H. Centoly, B. Stamford, 1982; W. Frans,
G. Gaisl, 1983; J. Henritse et al., 1985; S. Kumagi et al., 1982; E. Lopategui
et al., 1986; T. Mickelson, F. Hagerman, 1982; B. W. Smith et al., 1984). При
этом во многих исследованиях установлена высокая эффективность использования
нагрузок на уровне ПАНО. Такой режим тренировок значительно увеличивает порог
анаэробного обмена и максимальное потребление кислорода, снижает концентрацию
лактата в крови и ЧСС (V. Bung, 1983; H. Centoly, B. Stamford, 1982; W. Frans,
G. Gaisl, 1983; J. Henritse et al., 1985; T. Yoshida et al., 1982).
В эксперименте, проведенном в течение трех недель на двух
группах высококвалифицированных спортсменок, специализирующихся в академической
гребле, одна из которых (опытная) выполняла двигательные действия силового
характера в небольшом темпе (8-10 движений/мин) на тренажерных устройствах и в
естественных условиях при концентрации лактата, соответствующей ПАНО, заметно
увеличилась мощность на уровне порога анаэробного обмена (прирост составил 15,2
% р <0,01) и специальная работоспособность (на 9,1% р <0,01) при
неизменном уровне МПК. Вторая группа использовала греблю в темпе 26-28
гребков/мин, уровень интенсивности которой незначительно превысил (лактат 4-5
мМоль/л) мощность на уровне ПАНО. Эта группа не добилась достоверного прироста
специальной работоспособности при выполнении 3-х минутного теста в работе до
предела (1986).
Результаты многих исследований свидетельствуют о
целесообразности тренировочных упражнений, которые не только нагружают мышцы,
но и избирательно воздействуют на разные типы мышечных волокон. Ранее было
показано, что непрерывная нагрузка с интенсивностью 50-80% Vo₂max наиболее эффективна
для повышения окислительных возможностей волокон I типа. В то время, как
интервальная тренировка с интенсивностью большей или равной Vo₂max, наиболее
эффективна для повышения окислительных возможностей волокон II типа (J.
Henriksson, J. S. Reitman, 1976). Эти данные подтверждают гипотезу о связи
порога анаэробного обмена с окислительной способностью мышц (J. Karlson, I.
Jacobs, 1981; *****sko et al., 1980, 1986).
Концепция порога анаэробного обмена введена в спортивную
практику в начале 1960-х годов (K. Wasserman, M. B. Hroy, 1964) однако,
несмотря на большое количество исследований по этой проблеме не существует пока
единой теоретической базы данной концепции (G. A. Brooks, 1985; J. A. Davis,
1985). Это связано, как мы полагаем, с разрозненным освещением отдельных
положений в большом количестве обзорных статей (1985; G. A. Brooks, 1985; J. A.
Davis, 1985; J. Karlson, I. Jacobs, 1981; K. Wasserman, 1984) и главным
образом-с отсутствием общепринятой методики определения ПАНО.
Анализ литературных данных позволяет утверждать, что
принятие в качестве параметров, характеризующих ПАНО при ступенчато
возрастающих нагрузках, таких отдельно взятых показателей, как легочная
вентиляция (Vᴇ), выделение углекислого газа (Vᴄᴏ₂), потребление кислорода (Vᴏ₂),
ЧСС неправомерно (E. F. Hughes et al., 1982; M. R. Shorten, C. Williams, 1982).
При этом установлено, что пороги анаэробного обмена, определяемые по данным Vᴇ
и концентрации лактата в крови не совпадают, а накопление лактата в мышцах
возрастает раньше, чем обнаруживаются оба порога анаэробного обмена (H. J.
Green et al.,1983). Кроме того, обнаружено, что прекращение тестовой работы
анаэробно-гликолитической мощности, выполняемой в обычных условиях однократно
или повторно на фоне предрабочего алкалоза или ацидоза, происходит при самых
различных концентрациях лактата (7-15 мМоль/л) и показателях pH крови
(7,28-7,00), но при одинаковой концентрации лактата в мышечной ткани (24-25
мМоль/л на кг сырого веса) (, 1984). Как освещалось ранее, также неправомерно
использовать для характеристики ПАНО стандартных для всех испытуемых величины
лактата в крови (W. Kinderman et al., 1979; H. Shibayama, H. Ebashi, 1980; I.
Jocobs et al., 1981; и др.,1985) или излишка буфферных оснований (ВЕ) (V. Bung
et al., 1983).
Были предложены способы определения порога анаэробного
обмена по динамике избыточного выделения СО₂ (Ехс СО₂) (1973; J. Skinner, T. Mc
Lellan, 1980), путем нахождения переломов лактатной кривой (1983; M.
Pessenhofer et al., 1981; J. Skinner, T. Mc Lellan, 1980) и кривой температуры
ядра тела (1982, 1983, 1984, 1987), в точке перегиба на графике зависимости Vᴄᴏ₂
от Vᴏ₂ при ступенчато увеличивающейся со скоростью 15 Вт/мин нагрузки до
отказа, которая по мнению авторов методики, точно соответствует порогу роста
концентрации лактата и снижения бикарбонатов в артериальной крови (W. L.
Beauer, K. Wasserman, D. J. Whipp, 1986). С точки зрения терморегуляции тело
человека можно представить состоящим из двух компонентов: внешнего-«оболочки» и
внутреннего - «ядра». Температура поверхностных тканей оболочки, как правило,
ниже температуры глубоких тканей ядра. Ядро - это внутренние органы человека,
включая головной мозг. Регуляторные механизмы стремятся поддерживать
температуру ядра тела в покое и при неизменной температуре внешней среды на
постоянном уровне. Через оболочку идёт теплообмен между ядром и окружающей
средой.
Для оценки температуры ядра следовало бы измерять
температуру в сердце, где смешивается кровь из различных участков тела. Однако
это по понятным причинам не делается. Внутренние органы имеют разную
температуру, самый «горячий» - печень, ее температура доходит до 38-40°С.
Постоянство температуры тела человека обеспечивается
сочетанием двух взаимосвязанных процессов - теплопродукции и теплоотдачи. Если
теплопродукция преобладает над теплоотдачей, температура тела повышается. В тех
случаях, когда образование тепла меньше теплоотдачи,
наблюдается снижение температуры тела.
Теплообразование для человека — важнейший способ поддержания
температуры тела. Непрерывность обменных процессов в организме сопровождается
образованием тепла. Энергия, выделяемая человеком в сутки, слагается из 3-х
величин: энергии основного обмена, энергии повышения обмена при приеме пищи и,
наконец, энергии, образующейся в
результате умственной и физической деятельности.
Повышение теплопродукции в естественных природных условиях
происходит в основном при физической работе. Количество тепла, выделяющееся при
этом, зависит от вида деятельности, интенсивности и продолжительности работы.
Главный регулятор теплопродукции — мышцы. При интенсивной физической нагрузке
они поставляют до 90% тепла. В нормальных условиях на долю мышц приходится
65—70% теплопродукции. Второй по значимости источник теплопродукции - печень и
пищеварительный тракт, они дают% тепла. Резкое повышение температуры тела
наблюдается в момент отказа от дальнейшего продолжения физической деятельности,
выполняемой на пределе функциональных возможностей человека.
В наших исследованиях (1984, 1987) было установлено, что
косвенно о мощности двигательных действий на уровне порога анаэробного обмена
можно судить при выполнении ступенчато-повышающейся нагрузки, разделенной
паузами отдыха по началу увеличения тимпанальной температуры - температуры
барабанной перепонки, измеряемой через наружный слуховой проход, а точнее, как
доказали многочисленные исследования, температуры базальных отделов головного
мозга, главным образом гипоталамуса, центра вегетативных функций и
терморегуляции в том числе (Z. Mariak et al., 2003 и др.). Совместно с
отслеживанием с помощью цифрового термометра сдвига тимпанальной температуры,
определялся уровень лактата в капиллярной крови.
Весьма перспективным, по нашему мнению, является
использование инфракрасных тепловизоров в изучении воздействия различных
нагрузок на организм спортсменов. Современные тепловизоры позволяют с высокой
точностью оперативно производить оценку состояния регуляторных систем
организма; визуализировать процессы напряжения или срыва механизмов
терморегуляции при исследовании глубинных структур организма человека;
исследовать в динамике соматические и психоэмоциональные расстройства при
воздействии стрессовых факторов; выявлять патологические состояния на
дозонологической (предболезненной) стадии, определять функциональное состояния
организма спортсмена и его адаптационного ресурса, индивидуализировать
тренировочный процесс спортсменов (, 2012).
Как считал К. Wasserman (1984), порог анаэробного обмена,
выраженный в единицах Vᴏ₂ не зависит от способа и длительности двигательной
деятельности и связан с участвующей в ней мышечной массой. Лактат,
продуцируемый работающими мышцами, диффундирует в омывающую их кровь. При этом
на величину расхождений концентрации лактата в мышцах и крови влияют
интенсивность и длительность нагрузки, скорость кровотока, сочетание времени
работы, интервалов и характера отдыха ( , 1986 и др.). В другой своей работе К.
Wasserman (1984) установил, что при выполнении физической нагрузки ниже ПАНО
время достижения устойчивого состояния по потреблению О₂ - 3 мин., по выведению
СО₂ и легочной вентиляции – 4 мин., при превышении ПАНО устойчивое состояние
наступает позже. (1986) убедительно доказал, что сокращение времени работы на ступени
является источником ошибки определения ПАНО при ускоренном тестировании. Вторым
источником ошибки является, по нашему мнению, отсутствие или недостаточная
длительность интервалов отдыха между ступенями нагрузки. Они необходимы для
выравнивания лактата в мышцах и крови и для уменьшения «следов» предшествующего
воздействия на организм спортсменов, которое в свою очередь влияет на величину
порога анаэробного обмена (A. Black et al., 1984).
По мнению и (1983), для корректного определения ПАНО
необходимо пользоваться графическим способом его выявления при построении
лактатной кривой. Пороговая мощность или скорость нагрузки соответствует при
этом точке пересечения линий регрессии, построенных по экспериментальным
данным, забор крови осуществляется на 3, 5, 8-10-й минутах каждого из
интервалов отдыха при выполнении не менее 4-х ступеней нагрузки. Отметим, что
лактатную кривую необходимо строить на полулогарифмической шкале.
В нашем исследовании ( и др., 1988) при велоэргометрическом
тестировании, проведенном на 6 высококвалифицированных спортсменах-гребцах
(I-МСМК, 3-МС, 2-КМС), было установлено совпадение мощности на уровне ПАНО по
показателям динамики избыточного выделения СО₂ (Ехс СО₂) и концентрации лактата
(определяемых графическим способом), которая колебалась в широком диапазоне от
1,2 до 6, 2 мМоль/л. Длительность каждой из четырех ступеней нагрузки
составляла 5 мин., интервалов отдыха - 10 мин. Задаваемая мощность третьей
ступени нагрузки находилась в зоне предполагаемого уровня порога анаэробного
обмена (1500 кгм/мин).
Обобщая литературные данные и материалы собственных
исследований, посвященных поиску и сопоставлению различных методов определения
порога анаэробного обмена, следует заключить, что только кинетика лактата крови
в интервалах отдыха продолжительностью 5-8мин. между ступенями нагрузки,
длительность которых должна быть не менее 4-5 мин., а также динамика среднего
значения излишка неметаболической двуокиси углерода в выдыхаемом воздухе (Ехс
СО₂), определяемого в конце каждой нагрузки, наиболее адекватно отражают
процессы аэробно-анаэробного перехода энергообеспечения двигательных действий.
Подводя итог и обобщая все ранее изложенные факты, можно
сделать следующие выводы:
1. Отсутствие единой теоретической базы концепции порога
анаэробного обмена связано, как мы полагаем, главным образом с отсутствием
общепринятой методики определения этого важного показателя функциональной
подготовленности спортсменов в циклических видах спорта с преимущественным
проявлением выносливости. Наиболее точно он диагностируется значениями лактата
крови и избыточного выделения двуокиси углерода в выдыхаемом воздухе при выполнении
ступенчато возрастающей нагрузки, разделенной паузами отдыха. Использование для
характеристики порога анаэробного обмена стандартных для всех испытуемых
показателей неправомерно.
2. Чем больше вклад аэробных процессов энергообеспечения в
спортивный результат, тем больше должна быть доля физических упражнений,
выполняемых на уровне порога анаэробного обмена. Однако, применительно к
управлению подготовкой спортсменов необходимо реализовать принцип оптимальности,
проявляющийся в выборе минимально необходимой нагрузки для достижения
максимального спортивного результата.
3. Исходя из принципа оптимальности и учитывая, что порог
анаэробного обмена является нижней границей интенсивности нагрузок, развивающих
аэробные возможности организма, первостепенной задачей в циклических видах
спорта с преимущественным проявлением выносливости является определение
минимально необходимого времени воздействия нагрузки на уровне порога
анаэробного обмена в одном занятии, в микро-, мезо - и макроциклах подготовки спортсменов.
4. Методика развития аэробной подготовки в видах спорта с
преимущественным проявлением выносливости может быть представлена так:
- интенсивность на уровне мощности (скорости) порога
анаэробного обмена;
- продолжительность от 5 до 20 мин., большая
продолжительность может привести к значительному закислению крови и мышц в
случае превышения заданной мощности;
- интервал отдыха от 2 до 10 мин. необходим для устранения
возможного закисления организма;
- максимальное количество повторений в тренировке
ограничивается запасами гликогена в активных мышцах (примерно 60–90 мин.
чистого времени тренировки);
- тренировка с максимальным объемом повторяется через 2–3
дня, то есть после ресинтеза гликогена в мышцах.
5. Оптимизация процесса подготовки спортсменов в циклических
видах спорта возможна при использовании метода лидирования по скорости
передвижения или мощности двигательных действий при контроле за изменениями
темпа и временной структуры движения. Этот метод позволяет оперативно следить
за внешними проявлениями процесса утомления, постепенно повышать интенсивность
работы в зависимости от состояния тренированности в каждом цикле подготовки.
Определять в зависимости от цели занятия оптимальные временные и количественные
характеристики нагрузки, выполняемой различными методами тренировки и, что
особенно важно, добиваться более равномерного прохождения дистанции. При этом
ориентиром скорости должна быть прежде всего средняя скорость планируемого
результата (в процентах от нее), выбираемая с учетом физиологических и
биохимических сдвигов в организме спортсменов, степени психологических
напряжений. Нижней границей интенсивности двигательных действий в видах спорта
с преимущественным проявлением выносливости, развивающих функциональные
возможности организма спортсменов, должны быть тренировочные нагрузки на
длинных отрезках, преодолеваемых со скоростью 80-90% от планируемой средней
соревновательной.
6. Практическое использование на протяжении нескольких лет метода
лидирования с катера средней скорости передвижения лодки за гребной цикл с
учетом ее на уровне порога анаэробного обмена спортсменов и в процентах от
планируемой соревновательной при подготовке сборных команд РСФСР по
академической гребле к крупнейшим соревнованиям доказало его высокую эффективность.
Автор надеется, что приведенные в данной работе данные
позволят тренерам более вдумчиво и творчески подходить к тренировочному
процессу и будет благодарен за критические замечания и предложения.
18 мая 2012 г.
Шамиль Кашафович Агеев